2 forskellige veje ved aktivering af komplement
Mange af komplementkomponenterne i omløb er funktionelt inaktive. Aktivering af en komplementkomponent fører til aktiveringen af en anden komplementkomponent.
Den aktiverede anden komponent virker på den tredje komplementkomponent; aktiveringen af andre komplementkomponenter fortsætter på denne sekventielle måde. Således aktiveres komplementsystemet gennem en sekventiel kaskadlignende mode (fx arrangere 9 sten ved siden af hinanden og skub den første sten. Den første sten falder over den 2. sten og skubber den anden sten til at falde. Den anden sten falder på den tredje sten, som i turfald falder på den fjerde sten, og så videre indtil den sidste niende sten falder.) (figur 10.1).

Fig. 10.1 A til C: Et eksempel for at beskrive den kaskadlignende måde af komplementaktivering.
(A) Ni sten er anbragt side om side. (B) Den første sten skubbes og den falder og skubber den anden sten. (C) Efterfald af den første sten resulterer i sidste ende i efteråret af den sidste niende sten
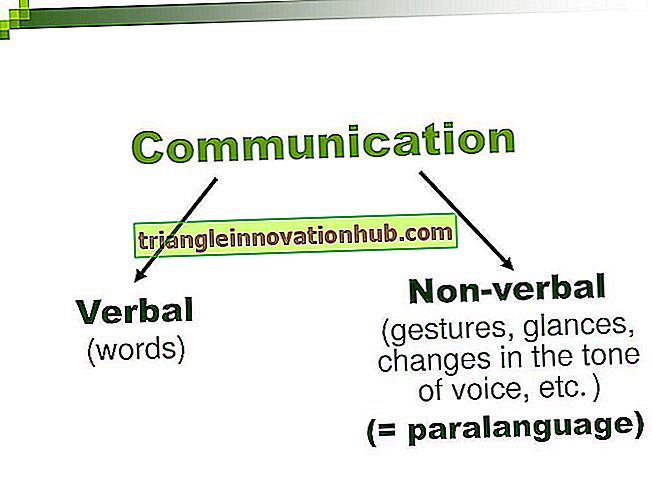
Aktivering af komplement sker via to forskellige veje:
jeg. Den komplementaktiveringsvej, der først blev opdaget, kaldes den klassiske vej for komplementaktivering.
ii. Den komplementaktiveringsvej, der opdages senere, kaldes alternativ vej for komplementaktivering. Mekanismerne for initiering af aktivering af disse to veje er forskellige. Alligevel fører begge veje til spaltningen af komplementkomponenten 3 (C3). C3 er fælles for begge veje, og de hændelser, der opstår efter spaltningen af C3 er ens i begge veje.
1. Klassisk vej af komplementaktivering:
Binding af antistof til antigen i omløb eller antigen på målcelle (såsom mikrobe) initierer aktiveringen af klassisk komplementvej (figur 10.2). Binding af antistoffer mod antigener udsætter C1q-bindingsstederne på Fc-regionen af antistofmolekyler. Komplementkomponenten Cl er fremstillet af 3 proteiner betegnet Clq, Clr og Clqr2S2. Clq-delen (af Cl) binder til C1q-bindingsstederne på antigenbundne antistoffer.
↓
Binding af C1q til antistof inducerer en konformationel ændring i C1r. Konformationsændringen gør C1r til et aktivt enzym, betegnet C1r.

Cir spalter C1s. De spaltede C1'er bliver et aktivt enzym designet C1s.
↓
C1s spinder igen to komplementkomponenter, C4 og C2.
jeg. C4 spaltes i C4a og C4b fragmenter. C4b-fragmentet er knyttet til den mikrobielle celleoverflade.
ii. C2-komponenten er knyttet til C4b. C2-komponenten bundet til C4b spaltes i C2a og C2b-fragmenter med C1s. C2b-fragmentet diffunderer væk, hvilket efterlader et C4b2a-kompleks på den mikrobielle celleoverflade.
↓
C4b2a-komplekset virker på C3 og spalter C3 i C3a- og C3b-fragmenter. (Eftersom den komplekse C4b2a spalter C3, kaldes C4b2a-komplekset også C3-konverterase.)
↓
C3b-fragment binder til C4b2a tørre former et C4b2a3b-kompleks. (Nogle C3b-fragmenter binder målcelleoverfladen og virker som opsonin for fagocytose af målcellen.
↓
C4b2a3b-komplekset spalter C5 i C5a og C5b. (Eftersom C4b2a3b spalter C5, kaldes C4b2a3b-komplekset klassisk pathway C5-konvertase.) C5b-fragmentet binder til overfladen af mikroben.
↓
C6 binder til C5b og danner et C5b6-kompleks.
↓
C7 binder til C5b6 og danner et C5b67-kompleks. Den hydrofobe region af C5b67-kompleks binder til phospholipider af den mikrobielle cellemembran, og C5b67-komplekset bliver indsat i den mikrobielle cellemembran.
↓
C8 binder til C5b67 og danner et C5b678-kompleks. C5b678-komplekset skaber en lille pore (10-A diameter) i den mikrobielle cellemembran.
↓
Mange molekyler (10 til 17 molekyler) C9 binder til en C5b678 til dannelse af C5b6789 (n) kompleks. C5b6789 (n) komplekset kaldes også som membranangrebskompleks (MAC). MAC-komplekset øger porestørrelsen til 70-100 A på den mikrobielle cellemembran (figur 10.3). Mange MAC'er dannes under komplement aktivering, og hver MAC er i stand til at punche et hul på cellemembranen (figur 10.4). På grund af det høje osmotiske tryk i mikrobialcellen kommer vand fra udvendigt ind i mikroben. Følgelig svulmer og spreder den mikrobielle celle (dvs. mikrobe lyserne).

Fig. 10.3: Membranangrebskompleks .
C5b6789 (n) komplekset dannet ved komplementaktivering kaldes også som membranangrebskompleks (MAC). MAC er et cylinderlignende produkt, og det slår et hul i cellemembranen. Gennem cylinderen som hulvæsker og molekyler strømmer ind og ud af cellen, hvilket resulterer i cellens død

Således resulterer aktiveringen af klassisk komplementvej med antigenbundet antistof i lysis af den mikrobielle celle, der udtrykker antigenet. Da den klassiske vej initieres af antistof, spiller den klassiske vej en rolle i erhvervede immunresponser. I mangel af specifikke antistoffer mod en mikrobe (som kommer ind i kroppen), vil den klassiske komplementvej ikke aktiveres (selv om alle komplementkomponenterne er til stede i kroppen).
De andre komplementfragmenter (såsom C4a, C3a og C5a) dannet under komplementaktiveringen har mange vigtige funktioner, og de beskrives senere (tabel 10.2).
Alternativ vej for komplementaktivering:
I modsætning til den klassiske komplementvej kræver den alternative komplementvej ikke antistoffer mod antigener til initiering af komplementaktivering. Dette indebærer, at den alternative vej aktiveres, selv under den første indtastning af antigenet. Med andre ord aktiveres den alternative pathway under et medfødt immunrespons. Den alternative komplementbane spiller en vigtig defensiv rolle mod mikrober, så snart mikroberne kommer ind i værten.
C3, faktor B, faktor D og properdin er de fire serumproteiner involveret i initieringen af alternativ komplementbaneaktivering (Figur 10.5, Tabel 10.3).
C3-molekylet har en ustabil thioesterbinding. På grund af thioesterbindelsens ustabile natur hydrolyserer C3 i blod spontant C3a og C3b. Hvis en mikrobe forekommer at være til stede nær C3b-dannelsen, føjes C3b-fragmentet til den mikrobielle celleoverflade.
↓
Faktor B binder til C3b på overfladen af mikroben.
↓
Faktor D virker enzymatisk på den C3b bundne faktor B for at producere to fragmenter, fragment Ba og fragment Bb. Fragment Ba diffunderer væk, og et C3bBb-kompleks dannes. C3bBb-komplekset har en halveringstid på kun 5 minutter. Men binding af et andet serumprotein kaldet properdin forlænger halveringstiden for C3bBb til 30 minutter.
↓
C3bBb-komplekset spalter et andet C3-molekyle til dannelse af C3a- og C3b-fragmenter. (C3bBb-komplekset hedder den alternative pathway C3-konverterase.) Fragmentet C3b binder til C3bBb og danner C3bBb3b-kompleks.
↓
C3bBb3b-komplekset spalter C5 i C5a og C5b (og derfor kaldes C3bBb3b som alternativ C5-konverterase). Efterfølgende komplementaktiveringstrin ligner trinene i den klassiske komplementaktiveringsvej.


C6 binder til C5b og former C5b6-kompleks.
↓
C7 binder til C5b6 og former C5b67-kompleks.
↓
C8 binder til C5b67 og danner C5b678-kompleks.
↓
Mange molekyler af C9 binder til C5b678 og danner C5b6789 (n) kompleks (membranangrebskompleks). Membranangrebskomplekserne punch huller i den mikrobielle cellevæg og fører til den mikrobielle lysis.

Amplifikation af komplementaktiveringstrin:
Komplementkomponenterne i blodet er i en funktionelt inaktiv tilstand. Nogle af komplementkomponenterne er proenzymer. Når proenzymet spaltes i to fragmenter, erhverver en af fragmenterne enzymatisk aktivitet.
Hvert enzymmolekyle dannet ved hvert trin af komplementaktivering virker på mange molekyler af den efterfølgende komplementkomponent, hvilket resulterer i aktiveringen af mange komplementkomponenter. Således øges antallet af komplementkomponenter aktiveret ved hvert trin mange gange, så der opnås et enormt antal membranangrebskomplekser og andre komplementfragmenter (f.eks. Kan et enkelt C3-konverterasmolekyle virke på 200 C3 molekyler og generere 200 C3b-fragmenter.) En sådan amplifikation af komplementaktivering ved forskellige trin hjælper værten med at producere et effektivt komplement-medieret forsvar.
Ikke-immunologisk aktivering af Classic Complement Pathway:
Normalt initieres den klassiske komplementbaneaktivering ved binding af C1q til antistoffet, der allerede er bundet til antigenet (dvs. initieringen af den klassiske vej for komplementaktivering er immunbaseret). Den aktivering af klassisk komplementbane kan imidlertid også initieres ved ikke-immunologiske midler.
jeg. Nogle bakterier (såsom Esch.coli og nogle Salmonella-stammer) og vira (såsom Para influenza virus og HIV) binder direkte til Clq og initierer komplementaktivering. En sådan ikke-immunologisk aktivering af den klassiske vej kan være nyttig for værten a) at reagere imod mikroberne som et medfødt immunrespons og b) at reagere imod mikroberne, selv før antistofferne binder til mikroberne.
ii. Uratkrystaller, bakterielle endotoksiner og heparin kan også initiere aktivering af klassisk komplementvej non-immunologisk.
2. Lectin-pathway of Complement Activation:
For nylig er en tredje vej af komplementaktivering kaldet "lectin-vejen for komplementaktivering" blevet beskrevet. Lectiner er proteiner, der binder til kulhydrater. Trinin-trinnets trin svarer til den for klassiske vej, bortset fra at lectinvejen ikke kræver antistof for at initiere komplementaktivering.
Mannosbindende lectin (MBL) er et akutfaseprotein produceret under akutte inflammatoriske responser. MBL har en struktur svarende til den af C1q. To andre molekyler kaldet MBL-associerede serinproteaser 1 og 2 (MASP-1 og MASP-2) er forbundet med MBL-aktiveringen af komplement-systemet. MBL og MBL-associerede serinproteaser antages at virke som C1q, C1r og C1s af den klassiske komplementvej.
MBL binder til kulhydratmolekyler på bakteriel celleoverflade.
↓
Derefter aktiveres MBL-associerede serinproteaser 1 og 2, hvilket fører til spaltningen af C4 og C2.
↓
De efterfølgende trin ligner dem i klassisk komplementaktivering.
Imidlertid er mange detaljer af lectinvej endnu ikke kendt. Da lectinvejen ikke kræver specifikke antistofmolekyler til aktivering, foreslås det, at lectinvej for komplementaktivering kan være en af de vigtige medfødte forsvarsmekanismer. MBL anerkender en bred vifte af klinisk signifikante bakterier, vira, svampe og parasitter.
Mutationer inden for MBL-genet er kendt for at resultere i suboptimale plasmaniveauer af MBL eller MBL-mangel. På grund af de overlappende virkninger af mange andre immunmekanismer forårsager lave niveauer af MBL normalt ikke kliniske symptomer hos immunkompetente personer. Men MBL-mangel er en signifikant risikofaktor for infektioner hos immunkompromitterede patienter (såsom kræftpatienter ved kemoterapi).